Эволюция сохранения и временной фрактал
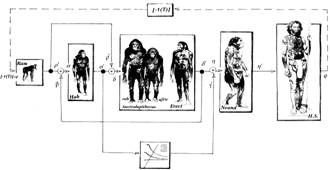 Отношение Времени и Пространства на клеточных множествах
Отношение Времени и Пространства на клеточных множествах
Анализ временных параметров модели управления качеством самосохранения показал некоторые особенности времени, связанные с вероятным существованием биологической системы его отсчета. Отношение кратности фрактала, как некоторого свойства сущности времени, проявилось как вторичное от первичных событий рецепции, поэтому, следуя теории Дж.Гибса и Х.Хармута, можно говорить, что событиями на клеточных ситуациях порождается не только время, но и отношения порождения собственного биологического времени системы самосохранения и "реального" времени.
Это означает, что в рассматриваемой ситуации, времени как к носителю объективной реальности процесса самоорганизации ракового гомеостата, могут быть свойственны дискретность, перетоки множества его квантов и цикличность. Если это предположение справедливо, эти отношения могут быть отображены граф-октавой времени как ситуационного носителя информации, т.е. объекта, состоящего из множества элементов, находящихся в ситуационных отношениях.
Рассмотрим с этих позиций в качестве ретроспективы, эволюцию Homo Sapiens как вида на отрезке времени в 10000 лет по земному календарю в Ньютоновской системе отсчета, связав его с полученным ранее отсчетом начала эволюционного процесса самосохранения. Как и раньше, предметом рассмотрения является оценка возможностей коррекции самосохранения, направленной на создание ракового гомеостата.
Это означает, что объектом-носителем формирующегося гомеостата мог стать вытеснивший неандертальского человека Neanderthal man кроманьонец Cro-Magnon man, по археологическим меркам современный человек, живший не более чем 25000 лет тому назад и образовавший, с его пещерным искусством и способами добывания огня, более прогрессивную ветвь эволюционного развития. Эта ветвь продолжена до сегодняшнего дня и не исключено, что современный цивилизованный Homo Sapiens сохранил до сих пор некоторые гены кроманьонца или даже неандертальца. Такие утверждения можно встретить в специальной литературе и они не противоречат тем временным параметрам процесса коррекции эволюции, которые мы обсуждали выше. Сохранение таких генов до настоящего времени может служить подтверждением незавершенности общего процесса эволюции человеческого рода как биологического вида. Напомним, что объектом эволюции является все многообразие функций, делающих человека сознательным элементом человеческой общности, как множества особей, связанных общими условиями, т.е. все то, что отличает человека цивилизованного от кроманьонца. Одной из компонент эволюции вида могла стать эволюция локального гомеостазиса и ролевой функции его собственного самосохранения.
Будем полагать, что общее число в 65 генетических изменений сохранения, с той или иной достоверностью, подтверждено нашими рассуждениями. Учтем также, что асимптотическим дополнением таких изменений, оказывается не 65-й, а 70-й виток. Этому не противоречит сохранение в Homo Sapiens отдельных генов, характерных для первобытного человека. Тогда результат простого деления временного интервала в 10000 лет на количество витков, равное 70 и соответствующее пределу, составляющему 13 циклов коррекции, согласуется с временным интервалом того отрезка эволюции, который мы назвали ее "витком". Он занимает ровно 142.857 (сто сорок два с лишним) условных года (года).
Последовательность ситуаций на вершинах графа для 70-го витка эволюции отображается полной серией указателей 999999 (табл.14.1). Это свидетельствует об асимптотическом завершении на этом витке технологического процесса коррекции. Если человек в настоящее время действительно находится на 65-м витке своей эволюции, ему предстоит пережить еще пять витков управляемой эволюции, т.е. еще 714.285 (семьсот четырнадцать с лишним) календарных лет. На этом эволюция сохранения завершается, локальный гомеостазис преобразуется в раковый гомеостат.
Как мы видим, длительность витка эволюции в принятых единицах времени в точности воспроизводит последовательность чисел 142857 модели, которая оцифровывает множество упорядоченных ситуаций на вершинах шестиугольника граф-октавы, начиная с седьмой вершины, замыкающей процесс. Из этого можно предположить, что:
- такое совпадение отражает на графе ситуативные свойства времени, определенного по Гибсу и Хармуту в пространстве Т-повторяемости, аналогично отражению и иных ситуативных объектов различной природы;
- в силу свойств граф-октавы как графа, отображающего ситуации на множествах, последовательность его вершин относительно объекта "Время", отображает не что иное как ситуации на множествах его квантов, находящихся в отношениях, которые сменяются циклически на фиксированных интервалах. При этом квантовые ситуации на вершинах графа образуются множествами квантов, порождающих различные функции многообразия, а характер порождаемых на этих множествах отношений, соответствует ролевым функциям клеточных множеств, взаимодействующих в групповых межклеточных отношениях;
- характер квантовых ситуаций на вершинах граф-октавы "Время" и образующих ситуационные отношения на них, остается неясным. Можно предположить, что сами ситуации и структурирующие их отношения, связаны с отношениями масштабирования и изменчивостью квантов времени, зависящей от ролевой функции сохраняемого на этих квантах клеточного множества;
- можно думать, что кванты времени, образующие ситуации, изменяемы и эта изменчивость связана с множествами структурных, динамических отношений, определяющих зависимость масштаба времени от динамичности сохранения ролевых функций клеточных множеств. Само Время, при этом, определяется перетоками квантовых ситуаций, образованных на множествах его квантов. Характер этих отношений остается неясным, но, скорее всего, это своеобразные отношения межквантовых взаимодействий. Их природу должны выяснять физики;
- в рамках развиваемых предположений можно говорить не только о дискретности времени, но и о двух его категориях:
- о кванте, как об образующем само Время элементе упорядоченного множества — структурной единице времени на квантовых ситуациях, генерирующей структурирующее процессы "Время", которое и можно полагать реальным, и
- о времени текущем — на перетоках множеств его квантов, структурирующем на молекулярном уровне исполнение события, порождаемого эволюционным процессом сохранения на клеточных множествах.
В этой связи возникает вопрос: какая категория времени масштабируется — текущее время или его структурная единица — квант. Представляется, что более динамична его структурная единица, построенная на динамических межквантовых взаимодействиях.
Итак, можно говорить, что временной фрактал, выявленный как специфическая особенность цикличности ситуативного объекта Время (по крайней мере для рассматриваемой ситуации формирования ракового гомеостата), кратен порядку, равному 10. Однако этот интервал связан с изучаемым объектом лишь внешней интерпретацией и никак не зависит от природы управляемой эволюции. Фрактал масштабирует время и зависит от его природы. Это — фрактал Времени. Его выявление, связанное с обсуждением проблем управляемой эволюции, чисто случайно. Можно предположить, что сущность последовательности 142857, в которой усматривают "магические" свойства, заключается в самоотображаемости ситуаций на множестве квантов Времени с фракталом 10.
Обсудим высказанные предположения более предметно. Для этого подвергнем сомнению календарное значение начала процесса формирования гомеостата. Допустим, что оно произошло не 10000 лет тому назад, а 100000 лет или даже 1000000 лет тому назад, или еще ранее — 10000000 лет тому назад. Рассмотрим, таким образом, числовую последовательность лет, кратную 10. Заметим, что последний член этого ряда 10000000 лет, по земному календарю, соответствует времени существования первобытной обезьяны Ramapithecus и периоду ее эволюционирования в Homo habilis. Антропологи считают, что именно Ramapithecus положила начало человеческой расе, о чем свидетельствуют и ископаемые останки. Этим значениям временного интервала соответствует последовательность временных интервалов витков 142.857, 1428.5714, 14285.714, 142857.1429 земных лет. Соответственно, оставшиеся пять витков управляемой эволюции, займут 714.285, 7142.857, 71428.571 и 714285.71 лет. Как видим, полученные значения кратны 10, а само число 10, рассматриваемое нами как дискретный временной инвариант управляемой эволюции, определяет такую кратность.
Полученные последовательности в трактовке смысла, приписываемого вершинам граф-октавы, отражают ситуации на них, свойственные Времени как объекту отображения Бытия. Это позволяет предположить, что полученное значение интервала времени с момента начала формирования ракового гомеостата отражает не календарное значение времени в единицах Земного календаря, а определенную ситуацию Времени, как носителя Бытия и оно равновероятно любой дате, кратной дискретному инварианту — фракталу управляемой эволюции, причем эта кратность отражает порядок (в смысле принадлежности временного интервала) некоторому ее фиксированному значению.
В любом случае, каждый из этих интервалов должен быть соотнесен к эпохам, связанным с данными современной археологической науки. Для уточнения конкретного временного интервала формирования локального гомеостазиса в раковый гомеостат, необходим сопоставительный анализ развития человеческой расы по виткам процесса управления эволюцией. Свойство цикличности вероятных календарных сроков начала эволюционного формирования ракового гомеостата, не позволяет уверенно подтвердить их полученное значение, хотя и не фальсифицирует факт его возникновения по принятой схеме самоорганизации.
Таким образом, мы пришли к характерной дробности размерности в пространстве — фрактальности. Это соответствует тому, что объекты клеточного мира самоподобны в пространстве. В данном случае мы, имея дело с сущностью времени, выявили дробность его масштаба, аналогичную дробности масштаба в пространстве. Это позволяет сделать высказывание — величина, названная дискретным инвариантом управляемой эволюции, отражает временную фрактальность дробности порядка десять этого процесса. Никакие другие значения дробности не приводят к аналогичному результату.
Из этого следует предположение, что если теория вторичности времени справедлива, то последовательности коррекций самосохранения (причина) и длительности корректируемой эволюции этих коррекций (следствие), являются "производными" от масштабного коэффициента — временного фрактала. Временной фрактал — понятие свойства Времени, отражающее с дробной размерностью понятие Пространства. Время и Пространство связаны отношением фрактала.
Итак, мы можем сделать некоторые выводы.
1. Для порождения и реализации функции самосохранения требуются определенные мощности клеточных компьютеров, ресурсы энергии, времени и пространства. Порождение функции самосохранения приводит либо к реструктуризации многообразия функций в ходе эволюции, либо к селективной коррекции отдельных механизмов сохранения, направленных на развитие избранных многообразий.
Коррекция многообразий в ходе порождения и эволюции функции самосохранения может быть ориентирована на повышение агрессивности клеток баланса эквивалентности, задания сагиттального направления роста или придания отдельным подмножествам клеток функций маркер-трассировщика или интенсификации этих функций.
Так как граф-модель отражает детерминированность всеобщего единства отношений пространства-времени и причина-следствие, не исключено, что она отражает также, и процесс эволюции всего живого мира. В частности, можно предположить, что такой граф отражает также процесс эволюции локального гомеостазиса в раковый гомеостат у ближайших предков человеческого рода. Завершение этого процесса, в сопутствующих условиях существования на измененной протяженности интервалов самокоррекции гомеостазиса, могло стать причиной гибели неандертальцев, без какого либо их противостояния с первобытным Homo sapiens.
Можно также сделать и более общее предположение, что граф-модель есть наиболее полное отображение самодостаточности клеточного множества, и обратно -идентификация с ее помощью любого ситуационного объекта, отражает его самодостаточность. Это высказывание, однако, требует специального рассмотрения и уточнения понятия "самодостаточности" клеточного множества, в первую очередь с позиций связи Времени и Пространства..
2. Промежуток времени с начала вековой коррекции эволюции сохранения, оказался достаточным лишь для ограниченных коррекций многообразий, не связанных с изменением класса функционирующих операторов. Такие коррекции возможны как на внутриситуационных перетоках, так и на транспорте клеток и их исполнение имеет характер непрерывных дискретных процессов на клеточных ситуациях. Витки коррекции формируют, вокруг оси спирали эволюции, спиральную "оболочку" переменного шага сложной конфигурации. Ее образующая зависит от характера управления, который задается дискретным временным инвариантом процесса.
Система самосохранения достигла качества, асимптотически близкого к качеству оптимальной системы, а система локального гомеостазиса — статуса, близкого к раковому гомеостату.
3. Витки оболочки и витки спирали существуют в разных временных пространствах. Отношения причинно-следственного характера между ними, отражают отношения между шагами и масштабом времени образующих их витков. Запуск и завершение процесса формирования ракового гомеостата реализованы между 15-м и 55-м витками коррекции, что соответствует 5 из 7 спиралей эволюции сохранения. Из общего количества 70 витков коррекции, 15 первых (1.5 спирали эволюции) образуют ее некорректируемый или слабо управляемый инициальный этап, а 5 ее незавершенных витков, соответствуют 0.5 спирали эволюции и отражают асимптотические переходные процессы достижения заданного качества. Интервал Т-повторяемости играет роль интервала управления.
Витки спирали эволюции, как и витки коррекции, характеризуются различной длительностью в пространстве реального времени.
4. Можно полагать, что самосохранение ролевой функции озлокачествления предполагает финитное значение некоторого "гарантированного остатка" клеточных множеств в 13%, не обладающего требуемым сохранением и развивающегося в режиме гомеостазиса. Не исключено, что 13% "гарантированного остатка" относится и к мощности подмножества особей человеческой популяции, имеющих шанс не подвергнуться атаке ракового гомеостата.
Вполне вероятно, что рост числа выявляемых сегодня злокачественных поражений отражает естественно-технологическое экспоненциальное повышение качества ракового гомеостата.
5. Процесс малигнизации эпителия и подлежащих тканей органов животных, в ряде случаев поддается управлению внешними агентами, что не свойственно органам человеческого организма. Химиотерапевтическое вмешательство после переключения гомеостазиса на локальный гомеостазис, обратить процесс не в состоянии. Функции ракового гомеостата человека — сохранение специфических отношений на клеточных множествах. Функция сохранения обуславливается развитостью межклеточных функциональных коммуникаций и возможностями обработки громадных объемов информации как на сетях распределенной клеточной компьютерной системы, так и на образующих эти сети клеточных компьютерах.
Раковый гомеостат реализует неотвратимо устойчивое сопротивление воздействию внешних агентов, направленных на изменение клеточных взаимодействий.
6. Анализ временных параметров модели управления качеством самосохранения, базирующейся на предположение вторичности Времени, выявил некоторые особенности его индикации, связанные с вероятным существованием биологической системы его отсчета. Последовательность вершин графа объекта "Время" отображает ситуации на множествах его квантов, порождающих отношения многообразия, которые сменяются циклически на фиксированных интервалах.
Характер порождаемых на этих множествах отношений, соответствует ролевым функциям клеточных множеств, взаимодействующих в групповых межклеточных отношениях.
7. Можно предположить, что ситуации на квантах времени и структурирующие их отношения, связаны с масштабированием и изменчивостью квантов, зависящей от ролевой функции сохраняемого на этих квантах клеточного множества. Отношения масштабирования образуются календарным генетическим фракталом, отражающим соотношение биологического времени коррекции и Ньютоновского времени эволюции. Эта величина, оказавшись постоянной, совпала в модели с основанием десятичной системы счисления. Само Время при этом, определяется перетоками квантовых ситуаций, образованных на множествах его квантов.
Временной фрактал — понятие свойства Времени, отражающее с дробной размерностью понятие Пространства. Время и Пространство связаны отношением фрактала.
8. В предположении вторичности времени естественно-технологического процесса эволюции сохранения, отражается существование двух его категорий. Квант — элемент упорядоченного множества, структурная единица времени на квантовых ситуациях. Он конфигурирует процессы "Время", в котором удобно создавать неподвижную систему отсчета и которое (по договоренности) можно полагать реальным. "Текущее время", в "подвижной" системе отсчета — переток множеств квантов, структурирующий исполнение события, порождаемого эволюционным процессом сохранения на клеточных множествах.
Квант — структурная единица времени, базирующаяся на межквантовых взаимодействиях. Не исключено, что именно она масштабируется в процессе сохранения.
9. Величина, отражающая дискретный инвариант управляемой эволюции сохранения, соответствует временной фрактальности с дробностью порядка десять этого процесса. Временной фрактал — специфическая особенность цикличности ситуативного объекта "Время", свойственная динамичности клеточной ситуации формирования ракового гомеостата. При условии, что теория вторичности времени справедлива, последовательности коррекций самосохранения и длительности корректируемой эволюции для этих коррекций, являются его флюксиями (производными).
Фрактал масштабирует биологическое Время и зависит от его природы.
10. Если полученные представления об управлениях на компьютерных биологических сетях достаточно реальны, то возможности самоусовершенствования локально гомеостазических управляющих программ до уровня ракового гомеостата свидетельствуют и об обратной допустимости — расшифровки программных кодов, с последующей заменой программ самосохранения на программы, разрушающие сложившийся раковый гомеостат и реверсирующие процессы сохранения.
Этому может способствовать изучение ракового гомеостата по данным клиники, использование отражающих его констант состояния — инвариантов в диагностике ранних состояний озлокачествления по персоналиям и изыскание диагностических и управляющих процедур реверсирования биологической программы ракового гомеостата в программу гомеостазиса.
Яков Гельфандбейн
(c) 1011494 CIPO
Популярное
Мария Шукшина поделилась постом, в котором высмеивается решение признать Ольгу Бузову лучшей телеведущей страны.



Последние материалы

"Правда.Ру" Эл № ФС77-72263 от 01.02.2018 года, выдано Роскомнадзором. Учредитель: Горшенин Вадим Валерьевич. Главный редактор: Новикова Инна Семеновна.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру».
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.